
Hello everyone, after moving to Rotterdam and having a baby (welcome Frank!) I am back here to show you the progress I made! The findings I present here are work in progress, but it is time to give a first update. I believe the results I present here are promising on three different levels: 1) content-wise, I think they are a start to understanding glycogen metabolism of Lactobacillus crispatus better 2) I am showing it all: data, methods, results, questions and flaws. Exactly how I envision Open Kitchen Science, (although with a bit of delay, so less “realtime” as I would like) 3) this is the work of a collaboration with another “postacademic” scientist, who wants to contibute to science besides her regular job. Meet Alicia Brandt!
Before diving into the science, a few points of order:
Collaboration: As mentioned before, I am very interested in collaborating and receiving feedback on my blog posts. Please contact me, or leave a remark. I am also interested whether any other labs are working on the same question or have been working on this in the past. If you are (or were ) doing experiments with Lactobacillus crispatus we could find out whether these results are true for more strains (especially using the same medium). Let’s talk! Twitter or Facebook or email rosanne dot hertzberger at gmail dot com
Manuscript: Part of the results presented here will be part of a paper that is currently in preparation by Charlotte van der Veer and Remco Kort and co-authors (including me). I will update this blog post as soon as this paper is published. When the paper is published, the full genomes of the strains will also be released online.
Internship: starting after the summer break I hope to be supervising a student working on this project, together with Jurgen Haanstra at the VU University Amsterdam. Find the description here (lab-site) or here (FigShare). BSc or MSc, please contact me if you are interested!
Warning: The results presented in this blog post are unreviewed and have not been replicated by an independent laboratory.
Open data All data and protocols used can be found in this FigShare collection.
How to cite: Hertzberger, Rosanne; Brandt, Alicia (2018): REBLAB: Carbohydrate active enzymes in Lactobacillus crispatus – a possible link between the pullulanase gene and growth on glycogen. figshare. Collection.
https://doi.org/10.6084/m9.figshare.c.4133819.v3
Short overview of where we are.
I am interested in glycogen metabolism of vaginal microbes. Glycogen is an abundant vaginal source of carbohydrates and varies depending on the vaginal bacterial signature. Roughly spoken: when the vaginal microbiota are dominated by Lactobacillus glycogen is generally higher. One of the overarching questions in the field is whether vaginal lactobacilli can metabolize this glycogen to grow and produce lactic acid. I think this is a pretty big deal. I have had a Google Scholar Alert with the keywords “vaginal glycogen” since a few years and not a week goes by without a paper getting published talking about how this vaginal glycogen is supposedly turned into lactic acid by lactobacilli. In my view, there is no evidence to back up this statement.
During my postdoc at Washington University St. Louis it became clear that the BV-associated Gardnerella vaginalis and Prevotella bivia are capable of growing on glycogen, as well as Lactobacillus iners (often encountered in vaginal microbiota with and without BV). Previously on this blog I showed experiments that Lactobacillus crispatus can grow and acidify using glycogen as a source. Here we zoom in further on the variation in glycogen metabolic capacity of different Lactobacillus crispatus strains, using a group of 23 different Lactobacillus crispatus isolates.
Summary of the findings
A dbCAN survey of the genomes of these strains shows an overview of several carbohydrate-active enzymes possibly involved in glycogen breakdown. One of the genes present in all strains is a putative S-layer linked type 1 pullulanase (GH13_13). Zooming in on this gene it appears that there is strong variation in the N-terminal sequence encoding a putative signal peptide. Only the strains that do not have a mutation in the sequence of this signal peptide show growth on glycogen. 6 out 23 strains that have a mutation in this peptide cannot grow on glycogen. Is this signal necessary for Lactobacillus crispatus to break down the big molecules of glycogen in the vaginal environment? Nothing final yet, but a promising lead to look further into.
Alicia Brandt: dbCan analysis and glycogen-active enzymes
Since I started with REBLAB I encountered several people who shared their own stories about their departure from science and how this affected them. Sometimes these people find the time and energy to still contribute to science parallel to their regular jobs. We start to call ourselves “post-academic” scientists. One of these people is dr. Alicia Brandt (previously Alicia Lammerts van Bueren as she is known in the glycobiology world) who left science last year for a job in a supporting role at the Young Academy at Groningen University. She expressed the strong wish to keep being involved and to share her knowledge and skill. It’s a happy coincidence that her expertise is exactly what I am looking for: glycobiology and even glycogen metabolism of bacteria. I went to meet her in Groningen and had a great time!

Alicia and me on the steps of the Groningen academy building
Lactobacillus crispatus isolates in this blog
Last year I was very lucky to join Prof. dr. Remco Kort at the VU University Amsterdam who had just isolated and sequenced about 30 Lactobacillus crispatus strains from women with and without BV ecology. These women were patients at the GGD facility in Amsterdam and the strains were isolated as part of the thesis work of Joke Dols. MSc student Jorne Swanenburg was responsible for the genomic analysis sofar and Charlotte van de Veer is currently finalizing her thesis and is writing up a paper on carbohydrate metabolism of these strains. Recently, a paper was published where the strains were used as a reference for a possible new vaginal prebiotic. The paper is open access and the isolation is described in materials&methods. At the moment we are analyzing these strains, some appear harder to culture than others. The list of isolates that we are able to maintain in the lab is a work in progress and will be updated later. We were unable to revive RL_005 from its glycerol stock and the sequencing file of RL_022 has some problems.
Alicia offered to use her expertise to help answer some questions surrounding Lactobacillus crispatus glycogen metabolism. First thing she did was to perform an analysis using the so-called dbCAN server, detecting the presence of several glycoproteins in the genomes of a set of isolated Lactobacillus crispatus strains that can potentially be involved in glycogen metabolism.
Find the raw data here (FigShare).
Find the protocol here (FigShare).
Most important findings:
- All L. crispatus genomes contained a putative cell-surface associated (as indicated by SLAP domain) pullulanase Type 1 enzyme (http://www.uniprot.org/uniprot/A0A135Z466) implicated in glycogen degradation. Key features of this enzyme are the presence of a CBM41 and CBM48 and a GH13_13 catalytic domain (Fig 1). More on this gene/protein later in this blog post.
- There is a cluster (operon?) of alpha-glucan degrading enzymes in all crispatus genomes which include a GH65 (phosphorylase?), GH13_20(CBM34) and GH13_31-2 enzyme. Further analysis required to see if they are co-transcribed. (Table 1, genes located in the cluster are indicated with an (a)).
- Other alpha-glucan metabolizing enzymes found within the L. crispatus genomes include: GH13_18, GH13_29, GH13_31, GH31 (see Table 1).
- Only three strains of L. crispatus contained a GH13_18 (RL02, RL09, RL10) (Note: RL_022 contained this enzyme as well, but sequencing file of strain has errors).
- RL_006 is the only strain that does not contain a GH13_29 enzyme.
Table 1: Overview of Alpha-glucan enzymes found in L. crispatus genomes (see www.cazy.org for more info on predicting enzyme activities based on amino acid sequence similarities with known enzymes within a given family)
| Enzyme Family | Proposed Activity | Genomes |
| GH13_13 (CBM41, CBM48) | Pullulanase type I | All except RL31, RL32 |
| GH13_18 | Sucrose phosphorylase | Only RL02, RL09, RL10. RL22* |
| GH13_20 (CBM34)(a) | Pullulanase type III, cyclodextrinase | All |
| GH13_29 | Trehalose-6-phosphate hydrolase | All except RL06 |
| GH13_31-1 | oligo-alpha-1,6-glucosidase | All |
| GH13_31-2 (a) | oligo-alpha-1,6-glucosidase | All |
| GH31-1 | alpha-glucosidase | All |
| GH31-2 | alpha-glucosidase | All |
| GH65 (a) | Maltose phosphorylase | All |
*RL22 genome needs to be resequenced (problems with sequencing file)
(a) Constitutes part of a cluster of enzymes, possibly an operon.
N-terminal signal peptide of the type 1 pullulanase gene (GH13_13) corresponds with growth on glycogen.
Glycogen is a prevalent potential carbon source in the vagina of reproductive age women. Lactobacillus crispatus and Lactobacillus iners-are the most frequently encountered species vaginally. They are generally assumed to be responsible for the low pH and high lactate concentrations- but it is unclear what sugar source they use for lactic acid production.
(To be fair: there are many assumptions and uncertainties here. To name a few: there is only circumstantial evidence that vaginal lactic acid is of bacterial origin. The same counts for the human origin of vaginal glycogen. Actually, the fact that we are dealing with glycogen and not with a different glycan is not all that well established since most studies used a PAS stain and an alpha-glucosidase. It could well be that we are dealing with a different glycan. However, I am cutting a few corners here and will assume that lactate is of bacterial origin and the glycan is in fact glycogen, produced by the hostess herself.)
Previously it was reported that at least a subset of vaginal lactobacilli (jensenii, gasseri and johnsonii) were unable to breakdown glycogen, but this study did not look at glycogen metabolism of crispatus and iners. I presented on this blog some evidence that the DSM strain of L crispatus is capable of growth and acidification (producing lactic acid) on glycogen as the carbon source.
This possible L. crispatus glycogen metabolism could be an important player in the acidification of the human vagina, and the health benefits that are associated with a vaginal community dominated by L. crispatus. Needless to say, we would like to know more about it.
Why the type 1 pullulanase had my interest – the role of serendipity
This gene had had my attention already since I started working with a set of 4 Lactobacillus crispatus strains during my postdoc in St Louis at the Lewis lab, WUSTL School of Medicine. These strains were MV-1A-US, MV-3A-US, JV-V01 and 125-2-CHN. I found that two of these (MV-1A-US and MV-3A-US) were able to grow on glycogen as a carbon source whereas two others (JV-V01 and 125-2-CHN) were not. To find a possible genetic origin of these differences I did a blastp analysis using several enzymes as a query that were known to be involved in glycogen (or starch) breakdown. Those were: glgX of E. coli, sap of Streptococcus agalactiae (see paper and sequence ), SusB of Bacteroides theta and the amylase (amyE) of Bacillus subtilis. Three out of the four genes showed no full-length copies in the four L. crispatus genomes, but I did find genes similar to the glgX gene: a gene annotated as a type 1 pullulanase (uniprot link of the copy in strain MV-3A-US). One of the strains unable to grow on glycogen (125-2-CHN) had no copy of this gene and the other strain unable to grow on glycogen (JV-V01) had a mutation in the upstream region, that I thought might be a dealbreaker for expression. This paper on comparative genomics of Lactobacillus crispatus confirms the absence/presence of the type 1 pullulanase in these strains. (supplementary material file nr 5). All other strains in the comparison contained the gene except for 125-2-CHN and 214-1.
This was all purely speculative at that moment, but this is why the type 1 pullulanase gene caught my attention: it was the only one that showed clear variation amongst the L. crispatus strains. There was a big chunk of serendipity that lead me to the presented finding.
All L. crispatus strains have the type 1 pullulanase gene, but the N-terminus looks different!
I was therefore disappointed that initially, from Alicia’s dbCAN analysis, and also in a screen performed by Jorne Swanenburg, it became clear that all strains had a copy of this gene. I further analysed this gene and also included the upstream region (expecting to find a mutation similar to the one in the JV-V01 strain). The sequence directly upstream this gene encodes a putative signal peptide (see Figure 1) and there is strong variation amongst the collection of genomes in this particular area (see Figure 2)
Please find the genes from all strains here. I used the EMBL Clustal Omega online Multiple Sequence Alignment tool to compare the genes and the results were striking. You could redo the analysis by using the file and just copy paste it into the Clustal Omega tool. The genes are conserved but not the starting region. In Figure 1, a schematic overview is shown of the organization of this gene in L. crispatus. Thanks Alicia!) In Figure 2, I am showing the variation in the N-terminal sequence from the aforementioned Clustal Omega comparison.
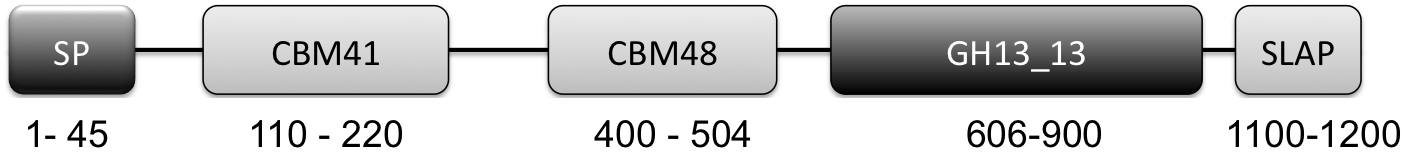
Figure 1: Graphical Representation of GH13_13 Pullulanase Type 1. (N to C terminal): SP: signal peptide (amino acids 1-45), CBM41: carbohydrate-binding module family 41 (amino acids 110-220), CBM48: carbohydrate-binding module family 48 (amino acids 400-505), GH13_13: glycoside hydrolase family 13 subfamily 13 (amino acids 606-900), SLAP: Surface layer associated domain (1100-1259)
 Figure 2: Comparison of the N-terminal sequence of the pullulanase gene in L. crispatus strains. Adjusted from Clustal Omega. Red: strains with a possible disrupted N-terminal sequence and signal peptide. Blue: strains with an N-terminal sequence indicating an intact signal peptide. Find full sequences here.
Figure 2: Comparison of the N-terminal sequence of the pullulanase gene in L. crispatus strains. Adjusted from Clustal Omega. Red: strains with a possible disrupted N-terminal sequence and signal peptide. Blue: strains with an N-terminal sequence indicating an intact signal peptide. Find full sequences here.
7 out of the 24 strains (RL_#-strains) have a mutation in the N-terminal locus of the pullulanase gene, more specifically, in the sequence of the signal peptide. Those strains are indicated by red in Figure 2. Further experiments should indicate whether this means that the start site of these “red” strains lies more downstream from the start site in the “blue” strains.
Remarkably, there are five different variants of this gene locus present in those seven strains. For instance, strains RL_002 and RL_009 only show a deletion of two nucleotides (a frame shift), whereas strain RL_006 and RL_007 have a completely different sequence in this region.
Growth on glycogen of the L. crispatus strains.
I performed a very straightforward cultivation experiment using glycogen as a carbon source. Initially, I only used four isolates (RL_002, RL_003, RL_007 and RL_026). When these results were promising I started to screen all 23 strains . As a benchmark I used the DSM strain, which I previously showed is able to breakdown glycogen for growth and lactic acid production. I used the same methods as described in that blog post:
I inoculated the strains in regular NYCIII glucose and after ~72 hours of growth diluted them with NYCIII glycogen (final concentration 0,5%), water (as a negative control) and glucose dissolved in water (final concentration 0,5% as a positive control). After 48 hours I measured the optical density at 600 nm to determine growth on glycogen compared to growth on NYCIII without supplemented energy source and NYCIII glucose.
For 19 out of the 23 strains tested (we could not revive strain RL_005) I have either biological triplicates or duplicates, I am showing their results in Figure 3. I am still working on getting all data complete and aim to have at least two replicates of this experiment for each strain. In one biological replicate of strain RL_019 and one replicate of RL_006 results are very different from the others. I have no idea why and no ‘reason’ to exclude it. Not really sure what to do with this measurement at this point. All individual biological replicates are shown in the figure.
Note: I measured the cell density 10x diluted in PBS as well as undiluted. I am showing the diluted data in this blog post, since I do not have the undiluted culture measured on all dates for all strains. The difference between “growth” and “no growth” is more pronounced in the undiluted measurement since background absorption of the media is lower. I include an overview of the undiluted data in a separate tab of the excel sheet and the figure in the GraphPad Prism file. Other data that are in the file but not in the Figure shown here: the optical density in the positive (glucose) and negative control (water). Find the data here (.xls and the GraphPad Prism 7 file to generate the figure).
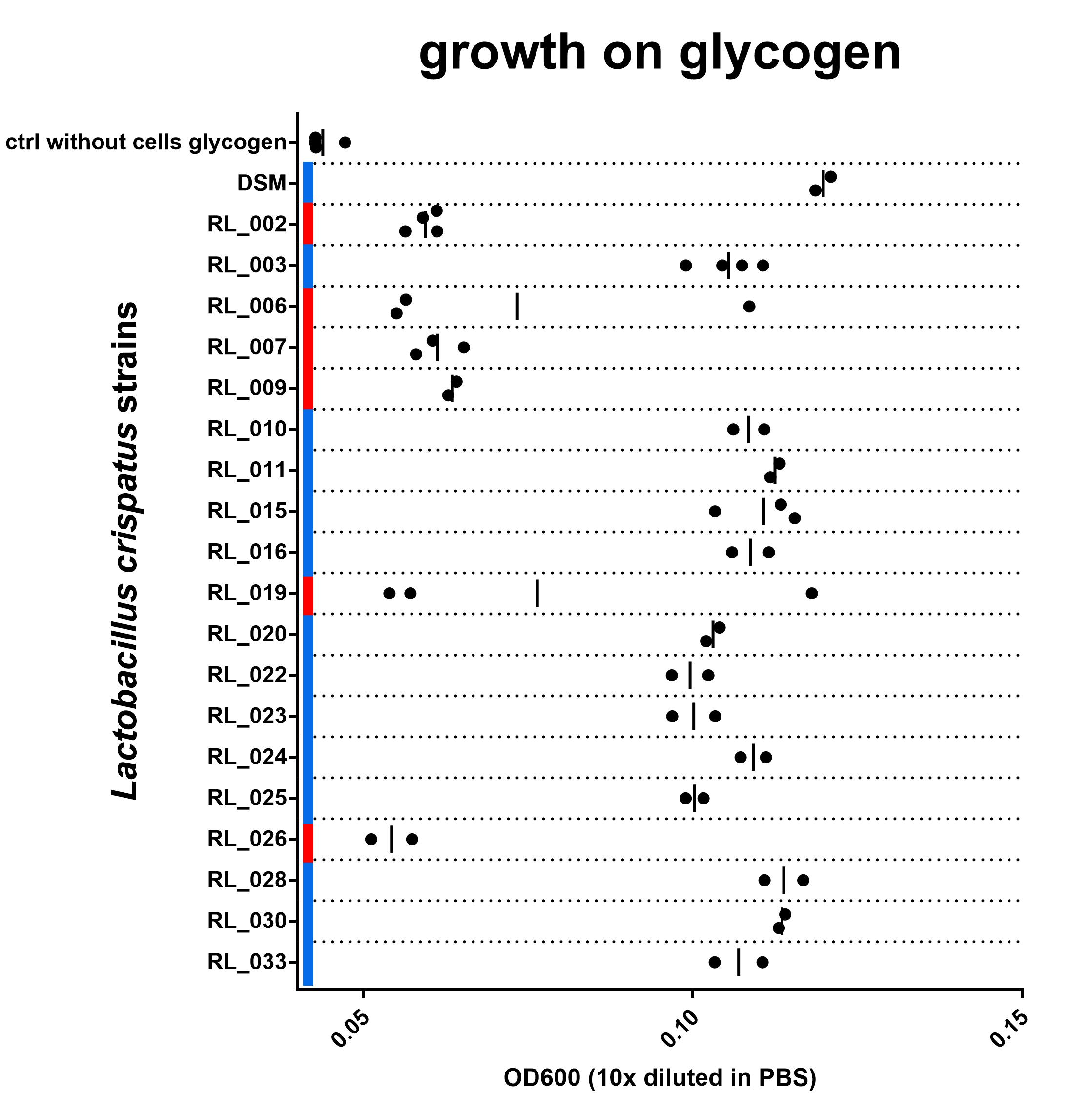
Figure 3: OD600 after culturing on NYCIIImedia with 0,5% glycogen. Biological replicates are shown as individual data points (some duplicates, some triplicates), vertical line indicates mean. Red or blue corresponds with red and blue in Figure 2. blue = with intact N-terminal signal peptide, red = with a disruption in this sequence. Find data here and protocol here
Although the growth data set is not complete yet, I do think we are seeing a strong connection between the N-terminal signal peptide of the type 1 pullulanase and the ability of the L. crispatus strain to grow on glycogen. All 6 strains in this experiment that have a disruption in the N-terminal signal peptide of the pullulanase gene show no growth on glycogen. The 14 strains that do have an intact signal peptide in the pullulanase gene can use glycogen as a source for growth. I see these data as strong evidence for an essential role of the pullulanase gene for glycogen consumption in Lactobacillus crispatus and, more specifically, the N-terminal signal peptide.
Thoughts, questions and new experimental plans
This finding is just that, a finding. Nothing final yet, an experiment that leads to a hypothesis: the N-terminal 29 amino acids are somehow important for Lactobacillus crispatus glycogen consumption. But how? Does this signal peptide lead to secretion of this enzyme? And if so, what is the influence of the C-terminal SLAP-domain? Are both required to localize this enzyme on the outer cell wall to be able to break down the big molecules of glycogen in its surroundings? Or does this signal peptide have a different function?
It is possible that the pullulanase without the signal peptide still functions in an intracellular metabolic pathway for glycogen breakdown, whereas the pullulanase gene that is localized on the outer cell wall can also debranch external glycans and utilize them for growth and lactic acid production.
I envision a few experiments to test these questions:
- it is probably important at this point to establish that the pullulanase is indeed a pullulanase. I have talked with Alicia about expressing the gene in E. coli to further characterize its activity. Alternatively, we could try to capture its native activity using cell free extracts and analyse carbohydrate products with Thin Layer Chromatography. Previously, we were able to show starch breakdown, next we should take a better look at the actual breakdown products.
- How can we study the role of the signal peptide? This is not that straightforward. Optimally we would just make L. crispatus mutants with and without it and track the enzyme’s activity and location. However, I have not seen any Lactobacillus crispatus cloning anywhere and not looking forward to try to develop my own protocols to get these isolates genetically accessible. (if someone has an idea, let me know). I hope to come up this summer with some experimental plan to test the cellular localization of this enzyme with and without the signal peptide.
What is the role of Lactobacillus crispatus glycogen metabolism in the context of the vaginal environment?
I believe that these experiments show that this activity is not something essential: the non-glycogen consumers seem to live happily in the vagina. Other lactobacilli, such as jensenii and gasseri also flourish in the vagina without being able to break down glycogen. Either, these bacteria utilize an alternative energy and carbon source. Or -what I personally expect- is that these bacteria live alongside glycogen consumers such as Lactobacillus iners or crispatus strains. Currently we don’t know whether the lactobacilli we encounter so abundantly in the vagina of reproductive age women are a collection of various strains or are a clonal population. It could well be that the non-glycogen consumers only thrive as freeloaders and can only colonize alongside a second glycogen-consuming species that does some of the glycogen breakdown. These exciting questions are definitely on my experimental wish list.
As I am still at home with baby, I will mostly do some reading and computer work to understand the genetic context and role of this pullulanase in other species. I found some interesting literature on this enzyme in Lactobacillus acidophilus and in a thermophilic bacterium called “Caldicellulosiruptor kronotskyensisencodes” (WOW! That must be one really interesting species!) Generally, pullulanases (and definitely secreted pullulanases) are of industrial relevance so there should be quite some protocols and knowledge out there. Also, I really need to read up on S-layers in lactic acid bacteria. Stay tuned!
And again: if you have suggestions how to continue, remarks or criticism, please let me know below. In general, I really appreciate any signs that this work matters to anyone because of the alternative publication route I am taking here.